
The debate over how people first arrived in the Western Hemisphere continues to roil archaeology in the United States – and to capture public attention. Today, the scientific community is contending with significant amounts of new genetic and archaeological data, and it can be overwhelming and even contradictory. These data are coming from new archaeological excavations but also from the application of newly developed tools to re-analyse prior sites and artefacts. They’re coming from newly sequenced genomes from ancient peoples and their contemporary descendants, but also from re-analysis of prior sequence data using new modelling tools. The generation of new data at times feels as though it’s outpacing efforts to integrate it into coherent and testable models.
Did humans first populate the Americas 100,000 years ago, 30,000 years ago, 15,000 years ago, or 13,000 years ago? Did they come by boat or by an overland route? Were the ancestors of Native Americans from one population or several? The answers to these questions would help us understand the grand story of human evolution. We know that the Americas were the last continents that anatomically modern Homo sapiens – humans like us – entered, but we don’t know exactly how this happened. These long-ago movements give us hints about the challenges ancient peoples across the world had to contend with during the Last Glacial Maximum (LGM), a prolonged period of coldness and aridity, when animals, plants and humans retreated to environmental ‘refugia’ for several thousand years. How did we survive this Ice Age? What technological and biological adaptations arose as the result of these environmental conditions? These questions capture the popular imagination and challenge the scientists working to uncover the details of individual lives thousands of years in the past.
To their Indigenous descendants, the stories we tell about these First Peoples of the Americas are highly relevant for additional reasons. Their deep ties and claims to the lands have often been ignored or expunged by governments, media and corporations across North and South America in order to make room for narratives that are more palatable, exciting or convenient to certain non-Native groups. The historical exclusion of Indigenous peoples from making decisions about research on their own ancestors and lands has caused significant harms to Native communities and individuals; when Native scientists and community members are full participants in the research process, the stories that emerge are not only more respectful but also more accurate.
Archaeological evidence establishes that Indigenous peoples were present in the Americas at least 15,000 years ago. Scientists don’t agree, however, on when people first arrived. Some archaeologists claim it must have been much, much farther back, citing evidence such as flaked stones in layers dating to ~30,000 years ago at the Chiquihuite Cave site in Mexico, bones with cut marks in layers dating to 34,000 years ago in Uruguay, flaked stones in layers dating to 30,000-50,000 years ago in Brazil, and even broken mastodon bones dating to 130,000 years ago in California. All of these claims are heavily disputed.

A general view of excavations at the White Sands site in New Mexico
As a rule, an archaeological site won’t gain widespread acceptance as legitimate unless there is clear evidence of human activity, that evidence can be securely dated, and it is found in an undisturbed geological context. For example, a hearth containing the remains of charred animal bone fragments and stone tool fragments at the Dry Creek site in Eastern Beringia (near the present-day Denali National Park in Alaska) was dated to 13,485-13,365 years ago from wood charcoal pieces taken from within the hearth. The stone tools – resharpened blades, flakes, end scrapers, and the byproducts of manufacturing them – and repeated controlled fires used to cook animal bones clearly indicate a human presence. The intact stratigraphy and multiple independent radiocarbon dates from the hearth tell us when people were using this particular part of the site. To archaeologists, this is uncontroversial. In contrast to the Dry Creek site, there is no consensus that the very early sites discussed above have met that standard; critics argue that the stone ‘artefacts’ and ‘butchering’ marks could be the result of natural phenomena (or even, in some cases, left by modern construction equipment). There simply hasn’t been any uncontroversial physical evidence of a human presence in the Americas more than 15,500 years ago.
As they walked along the muddy surface, their feet mushed tiny seeds of ditch grass into the ground
Then, in 2021, a team of archaeologists dropped a bombshell into this debate: they’d found footprints – unquestionable evidence of a human presence – at White Sands National Park in New Mexico, dating to between 23,000-21,000 years ago.

Footprints from Site 2 at White Sands
The White Sands Locality 2 site was once the shore of an ancient lake. For more than 2,000 years, humans and animals visited it. As they walked along the muddy surface, their feet mushed tiny seeds of ditch grass into the ground, leaving a vital organic trace that archaeologists can use for carbon dating. (Some archaeologists have criticised the dating methods used, but there is general agreement that the presence of human tracks with fauna known to have gone extinct around 11,000 years ago dates these to – at minimum – the end of the Pleistocene.) If the find holds up to scrutiny, physical evidence of a human presence in the Americas during the LGM would be a paradigm-changing event, pushing back the date of the earliest migrations to sometime before 25,000 years ago.
When European settlers and explorers first encountered the Native peoples of the Americas, they sought to force the fact of the Native people’s existence into a Biblical worldview. The First Peoples, who built the impressive earthworks, monuments, temples and pyramids throughout the Americas, were recast as members of a lost tribe of Israelites, Irish sailors, or possibly Vikings, for the ideological convenience of settlers.
The pretence that the first peoples of the Americas were a different race than Native Americans – a view known today as the Myth of the Moundbuilders – became extremely popular in the 18th and 19th centuries. Andrew Jackson explicitly used it in 1830 to justify the brutal Indian Removal Act:
In the monuments and fortresses of an unknown people, spread over the extensive regions of the West, we behold the memorials of a once powerful race, which was exterminated or has disappeared to make room for the existing savage tribes.
As the field of archaeology matured and incorporated the scientific method, scholars began to reject the Moundbuilder Myth. By the end of the 19th century, the US government funded an investigation of mounds throughout North America to identify their creators. The evidence persuaded researchers that the mounds were built by the ancestors of contemporary Indigenous Americans, not some mysterious, lost race. The resulting Twelfth Annual Report of the Bureau of Ethnology to the Secretary of the Smithsonian Institution (1890-91) marked a new era in archaeology. In time, the archaeological, cultural and biological evidence all pointed to shared ancestry with Asians, suggesting that the ancestors of Native Americans came to the continents via a land bridge between Siberia and Alaska.
The question of when did this migration begin remained. Poorly understood geological and cultural chronologies made it a difficult matter to address. Radiometric dating methods were not invented until 1946, and strong rivalries between scientists promoting their own models confused the issue.
In the Clovis First model, hunter-gatherers travelled from Siberia to North America across the Bering Land Bridge
The 1927 discovery of a human-made projectile (spear point) at the Folsom site in New Mexico proved a turning point. The spear point was associated with the remains of a bison that had gone extinct at the end of the Pleistocene (around 11,700 years ago). Even without radiometric dating methods, researchers knew that a human artefact embedded in an animal that went extinct long ago had pushed back the date at which humans were living in the Americas. It marked another important shift in the study of human origins in the Western Hemisphere. Archaeologists used such artefacts to begin piecing together a bigger historical narrative.
The model that emerged and dominated the field for decades has come to be called ‘Clovis First’. It is named after the technologies first found in 1929 at a Pleistocene kill site near the town of Clovis in New Mexico. Characterised by a thin, lance-shaped projectile point flaked on both sides with a single, long flake removed at its base on each side (known as a ‘flute’), the Clovis point is unique to North America. It appeared widely across the continent beginning about 13,400 years ago, near the end of the LGM.
Because of their sudden appearance and spread, and in the absence of recognised earlier sites, many archaeologists believed that the Clovis tools were evidence of the very earliest inhabitants of the Americas. The Clovis First model held that a small group of hunter-gatherers travelled from Siberia to North America across the Bering Land Bridge. They then followed southward an ice-free corridor that warming temperatures had opened along the eastern Canadian Rocky Mountains. The population dispersed rapidly across the continents, leaving their newly invented projectile points embedded in their prey, or cached in special locations, along the way.
The Clovis First model faced challenges from growing evidence of an earlier human presence in the Americas. Some of it was archaeological; signs of a human presence at the Monte Verde site in Chile dated to more than 1,000 years before the first appearance of Clovis. As the science of genetics matured, it added a critical source of evidence against the Clovis First model. Sequencing of DNA, and assuming that DNA bases mutate at a known and constant rate, allowed geneticists to estimate when different populations last shared a common ancestor. Depending on which mutation rate was used, the genetic evidence suggested a last common ancestor for Native Americans anywhere from 15,000 to 30,000 years ago, or 2,000 to 17,000 years earlier than accounted for by the Clovis First model.
This is not all that’s revealed by genetic studies of mitochondrial DNA from ancient and contemporary Native Americans. They also showed that the ancestors of the First Peoples had undergone a period of isolation before their lineages diversified. The molecular clock – a dating method based on the rate at which DNA mutates – tied this isolation period to roughly the height of the LGM, beginning about 21,000-20,000 years ago. This is when climate conditions across the globe forced people and animals to retreat into refugia to escape the encroaching ice. When, 17,000 years ago, a potential coastal route in the Americas opened as the ice sheet retreated from the Pacific, lineages began to diversify more rapidly as they spread across geographic space and encountered each other less frequently to spread newly arising genetic variants.
In the last generation, geneticists have integrated their data with archaeological and climatic evidence to produce a three-stage model for the peopling of the Americas. First, approximately 20,000 years ago, a group of Asians migrated into Beringia. They remained isolated there for thousands of years, during which they evolved the founding lineages seen across the Americas. Then about 17,000-16,000 years ago they migrated out of Beringia into the Americas, rapidly peopling the continents. Still later, migrations from Beringian populations then peopled the Arctic. The genetic variation present in Indigenous peoples of the Americas was the result of local evolutionary processes with no gene flow from populations outside the continents. This mitochondrial DNA-based model has come to be known as the ‘Beringian standstill’ or ‘Beringian incubation’ hypothesis.
Native Americans today can trace their genomes to Ancient North Eurasians and an ancient East Asian group
The genetic evidence for the Beringian standstill hypothesis was based on certain types of DNA – like mitochondrial DNA and Y-chromosome DNA – that are inherited from just one parent. These are relatively easy to extract, work on and understand, but they have limitations. The full human genome is massive, with 3 billion base pairs and some 20,000 genes. Each stretch of DNA has its own history of evolution, and each may closely resemble that of the parent population or differ from it in important ways. The most accurate histories that can be reconstructed from DNA come from the study of the entire genome, not single genes.
Getting complete genomes from ancient individuals is difficult. After an organism dies, the cellular processes responsible for repairing damage to DNA cease to work. Over time, an organism’s DNA fragments and accumulates damage until at some point it is no longer possible to recover; exactly how quickly this occurs depends on a combination of environmental conditions and time. In 2010, Danish and Chinese scientists led a study that mapped the first complete genome of an ancient human, from a Palaeo-Inuit Saqqaq man who lived 4,000 years ago in Greenland. This work from Morten Rasmussen, Yingrui Li and Stinus Lindgreen brought the ‘palaeogenomics revolution’ to the Western Hemisphere and transformed our understanding of human history across the continents.
Complete genomes from ancient humans in the Americas allowed researchers to model biological histories on a scale never imagined before. Because each nuclear genome reflects the contribution of thousands of ancestors, entire population histories can be reconstructed from just a few individual genomes. Genomes from the past can often reveal details of human biological history that were obscured by later demographic events (such as population migrations). While models generated from genomic data can’t give insights into all aspects of human history (particularly issues pertaining to cultural identity), they nevertheless provide a powerful way of understanding biological relationships through time.
The sequencing of ancient genomes in Siberia brought one of the most important insights into Native American population history. The first evidence of humans living above the Arctic Circle comes from the Yana Rhinoceros Horn site in northeastern Siberia, where year-round settlements of humans have been dated to 31,600 years ago. From DNA retrieved from the baby teeth of two young children growing up at the site, geneticists were able to piece together a picture of a large Upper Palaeolithic population ancestral to the Ancient North Siberians, a group identified from the genome of a child buried at the Mal’ta site near Lake Baikal 24,000 years ago. The Ancient North Siberians were ancestral to Siberians, Central Asians and Europeans. The genome from the Mal’ta child revealed to palaeogeneticists that this group of Ancient North Siberians contributed some ancestry to the Beringian population who gave rise to the First Peoples. Native Americans today can trace between 14-38 per cent of their genomes to the Ancient North Eurasians, with the rest from an ancient East Asian group, likely from China. This gene-flow event between the First Peoples’ ancestors took place sometime between about 23,000-18,000 years ago.
Population models based on ancient genomes confirmed the Beringian standstill hypothesis. This means that the population’s isolation coincided with the height of the LGM, approximately 23,000-19,000 years ago, when sea levels would have been approximately 100 metres lower than today, and water bound in continental ice sheets. Across Siberia, as in other parts of the Northern Hemisphere, conditions would have been extremely dry and cold. In fact, the archaeological record indicates that Siberia was essentially depopulated. Genomes cannot tell us where the ancestors of Native Americans were isolated but they do tell us that they were isolated. Many geneticists believe that, because they were isolated, it’s unlikely they were living in East Asia during the LGM – there were other groups in the region, and proximity would likely have resulted in gene flow. The southern coast of the Bering Land Bridge in central Beringia is a likely candidate for a refugium, as palaeoclimactic reconstructions show us that it had a relatively mild climate, with abundant plant and animal resources. Since what was the Bering Land Bridge now lies mostly under the Chukchi and Bering Seas, it is difficult to test this hypothesis. Some archaeologists are investigating potential LGM-period sites with hints of human presence at Bluefish Caves in the Yukon, or at Lake E5 in the Brooks Range of Alaska. So far, no definitive evidence of a human presence during the LGM has been found in western Beringia.
Since we cannot tie the population ancestral to the First Peoples to any specific technological or cultural manifestation in the archaeological record, their location during the LGM between 23,000 and about 16,000 years ago remains a mystery for now.
One of the more astonishing insights palaeogenomes have given us is that the population ancestral to Native Americans split into several branches during its isolation between approximately 24,000 and 16,000 years ago. Geneticists refer to one of the branches as the ‘Ancient Beringians’. The Ancient Beringians were identified through the genomes from children buried 11,500 years ago at the Upward Sun River site in the Tanana River Valley in Alaska and from a young girl buried 9,000 years ago at the Trail Creek Cave site on the Seward Peninsula. These genomes suggest that the Ancient Beringian population was widespread across what is now Alaska, but also limited to Alaska. It did not persist (genetically) into the present day; Indigenous Arctic populations do not seem to have Ancient Beringian ancestry. We may eventually find people with Ancient Beringian ancestry as we characterise more genetic variation in present-day and ancient people, or it may be that this population ultimately diminished without leaving any descendants.
Another branch emerged approximately 24,700 years ago. ‘Unsampled Population A’ was identified indirectly from its contribution to the genomes of the ancestors of contemporary Mesoamericans and South Americans.
The First Peoples’ migration was mainly by boat along the west coast of Alaska, rather than on foot
The third known branch that emerged during the isolation period is known as ‘Ancestral Native Americans’. It is ancestral to all populations south of Alaska. Its dispersal southward throughout the Americas, most likely down the Pacific coast, saw a series of population splits. The earliest of these populations, identified by the genome of an individual from the Big Bar Lake site in British Columbia, split from the other Ancestral Native Americans as early as 21,000 years ago. The other two groups, known as ‘Northern Native Americans’ (in Alaska and northwestern Canada) and ‘Southern Native Americans’ (south of Alaska and Canada), diverged from each other some time after 17,000 years ago as they moved south.
Southern Native American populations dispersed throughout the Americas extremely quickly. We know this from the rapid divergence of lineages in ancient members of these groups. Because the speed and timing of their divergence is earlier than the most likely date for the opening of an interior route through the glacial ice (the Ice-Free Corridor along the Eastern Canadian Rockies), the model suggests that the First Peoples’ migration was mainly by boat along the west coast of Alaska, rather than on foot. In fact, the earliest archaeological evidence of people in the Ice-Free Corridor dates to shortly after 13,000 years ago, and indicates they were moving northward, rather than southward.
Following the initial dispersal sometime after about 21,000 years ago, the next 20,000 years brought a complicated series of population movements throughout South and North America. Glaciers that had been preventing eastward movement across the North American Arctic melted, and two successive migrations then peopled the eastern half of the continent. The first was by Palaeo-Inuit around 5,000 years ago, and the second by the ancestors of contemporary Inuit peoples between about 1,000 and 750 years ago. These two groups were culturally very different, but shared ancestry with Siberian populations, and at least the direct ancestors of living Indigenous Arctic peoples shared gene flow with members of the Northern Native American group. Population geneticists are still trying to sort out the complicated population histories of different Indigenous groups within the Arctic from both contemporary and ancestral genomes.
On a Saturday afternoon in April 2022, I sat in a packed hotel conference room in Chicago at the annual meeting of the Society for American Archaeology. It’s not a conference I usually attend, but I’d been asked to serve as a discussant in a session presenting the results from the White Sands site in New Mexico. I had decided in advance that my presentation would focus on trying to reconcile the genetics and archaeological records.
As I sat squeezed in between Dennis O’Rourke, the anthropological geneticist who was serving as the other discussant, and E James Dixon, an eminent archaeologist, I was excited to hear about the excavation of the White Sands site directly from the archaeologists who had studied the footprints up close. To a rapt audience of archaeologists – which included many sceptics of its purported 23,000-21,000-year dates – Vance Holliday, Thomas Urban, Clare Connelly, David Bustos, Amber Kalush, Matthew Bennett and Daniel Odess discussed their excavation of the site, how they obtained dates, how they worked with tribal partners, the future of the site, and the implications of the dates in the context of current models for the peopling of the Americas.
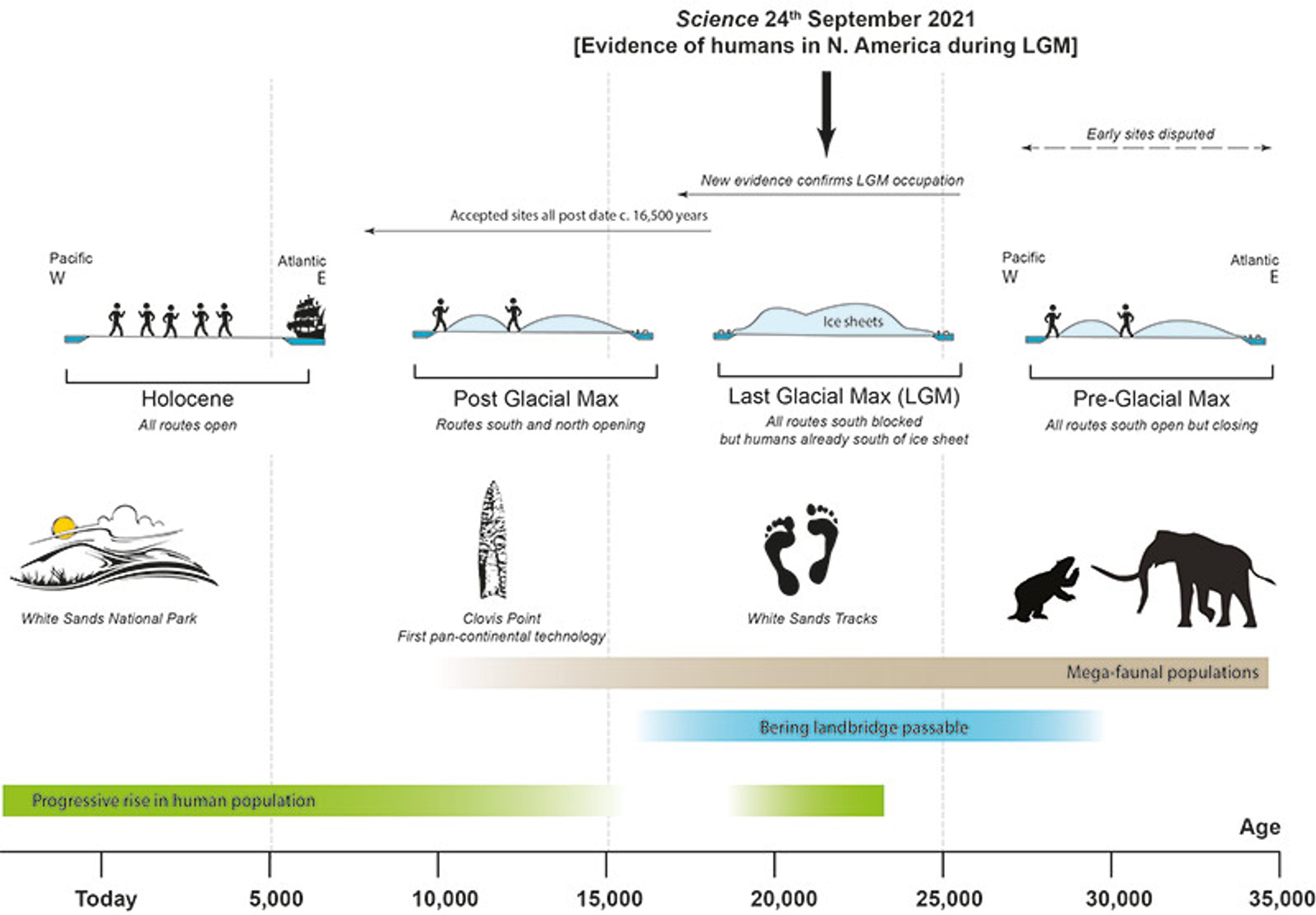
Timeline relating to the discovery of the tracks at White Sands
They had a formidable task; White Sands presents a radical challenge to our understanding of the earliest humans in the continents. If the 23,000-21,000-year dates are accurate, it would mean that people had to have made their way into the continents before the ice sheets fused, by 25,000 years ago or earlier. What could account for the huge time gap between the White Sands site and the archaeological and genetic evidence showing a migration after 17,000 years ago?
The seeds embedded in the footprints were not from plants growing in water but on the lakeshore
I recognised several people in the audience who had been shocked and sceptical when the dates had been published a few months prior. ‘They just can’t be right,’ an archaeologist told me at the time. There are some legitimate concerns about the site, which were raised during the question period of the symposium and in subsequent publications. These objections are not, as at other early sites, that the evidence for humans was ambiguous. There could be no question the footprints were human, and their immediate proximity to mastodons and giant sloths meant that they were unquestionably Late Pleistocene. Rather, the major concern raised primarily by geoarchaeologists is about the dating of the Ruppia cirrhosa seeds found embedded in the footprints.

The ditch grass seeds from which the team were able to date the excavations
Ruppia (ditch grass) can be influenced by the so-called hard-water effect in which plants growing in water take up old carbon present in it, making samples appear hundreds to thousands of years older than they actually are. Sceptical archaeologists believe that the hard-water effect may be biasing the carbon-14 dates, and that additional dating – ideally of features with other carbon sources – is needed to confirm the ages of the site.
Another concern, raised by the archaeologist C Vance Haynes of the University of Arizona, concerns the stratigraphic integrity of the sites. Specifically, he raised the possibility that the Ruppia seeds might not be from the same geological layers as the footprints. Rather, he suggests, the seeds might have been redeposited by wind from older strata elsewhere at the site into younger contexts. The apparent co-occurrence of human and Late Pleistocene animals at the site might be due to humans crossing over extinct animal trackways thousands of years later.
If this scenario is correct, additional dating from other organic sources (besides Ruppia seeds) should eventually demonstrate that the geological layers that contain the footprints date to after the LGM, perhaps to the Clovis period, which begins around 13,000 years ago.
But during the symposium and in a subsequent publication led by Jeffrey S Pigati of the US Geological Survey, the archaeologists who excavated White Sands robustly responded to these concerns. The hard-water effect was not biasing the dates, they explained, because the seeds were not from plants growing in water, but rather on the lakeshore, and because all dates were in good stratigraphic order. There was no possibility that the human footprints postdated the Late Pleistocene animals’ footprints because in many cases they were in layers underneath (and therefore geologically older than) animal trackways.
To each objection or concern, they have given a patient and convincing answer. And yet the debate rages on.
However frustrating this debate may be for all participants – and I imagine it can be very frustrating at times – to me it is an excellent example of the dynamic process of the scientific method. Claims are advanced, they are robustly criticised, and additional evidence is brought forward to refute the critiques. Bit by bit, the field advances in its understanding.
I sincerely hope that the public understands this debate, and indeed the broader debate about details regarding the peopling of the Americas, in this light. Unfortunately, because science is all too often taught in school as a collection of facts, rather than a dynamic process of enquiry, people can be vulnerable to being misled by opportunists such as Graham Hancock. A self-described investigative journalist, Hancock has made a career of weaving metaphysical stories about the past for audiences eager to believe that there is a conspiracy among thousands of archaeologists, geologists, palaeoclimatologists and geneticists like myself to suppress the ‘truth’ about the past. In his Netflix series Ancient Apocalypse (2022), Hancock fantasises about imaginary ‘alternative’ theories of the past, including the Americas, telling audiences they can acquire suppressed knowledge without the tedium of actually engaging with evidence.
But, as we have seen, debate about population history models is a normal part of the scientific process. Contrary to claims made about us, scientists emphatically don’t want dogma to stifle the consideration of new ideas. In contrast to ‘alternative historians’, scientists demand a rigorous evaluation of evidence in considering these ideas.
In contrast to metaphysical storytelling, we have testable hypotheses
There is certainly robust debate about the peopling of the Americas. White Sands, along with other pre-LGM sites, provides a good example of how this debate progresses. For most sceptical archaeologists, the attitude to the claims of the White Sands findings is not hostility, but a cautious ‘let’s wait and see’. It may be that, just as the Folsom and Clovis sites showed archaeologists where to look for evidence of Late Pleistocene humans associated with the remains of extinct animals, White Sands may show us where to look for even earlier peoples. Or it may be that additional dates will eventually show us that White Sands is younger than the LGM.
The important thing is that, in contrast to metaphysical storytelling, we have testable hypotheses. We know what evidence is needed to test these hypotheses, and archaeologists are out in the field right now gathering that evidence.
Outstanding questions
One of the limitations of our current understanding of biological population histories in the Americas is that they are based on very few ancient genomes. Our sampling of genetic diversity across the Americas is very incomplete. The reasons for this are in part historical. Non-Native scientists inherit a legacy of insensitive and exploitative research from our forebears, one that has left Indigenous peoples with little incentive to trust us with the remains of their ancestors. If we wish this to change, we have a great deal of work to do and many factors to consider. There are ongoing discussions about what constitutes appropriate ethical practices in our field, including who is doing the research, who is interpreting the results, and who has the right to determine what is done with the data that result from it. The histories we infer from archaeology and genetics are not abstract to contemporary Native Americans, who have their own scientific and historical knowledge stretching back for countless generations; how we tell the stories that emerge from our genetics data are of critical importance to them. The acquisition of DNA from ancient individuals usually requires the destruction of small portions of bone or teeth; this may be incompatible with a community’s values for how ancestors should be treated.
Any palaeogenomics project must contend with these issues, and that often takes a great deal of time. The slow nature of the consultation process – building relationships, respecting ‘no’ from communities who refuse research on their ancestors, co-designing a project with community partners from those who wish to do so – is at odds with the fast-paced, intensely competitive research environment characteristic of palaeogenomics. But if we wish to do research in a better way, this process cannot be rushed or circumvented in the pursuit of data.
Our limited sampling of ancient genomes may mean that there is still more genetic variation than accounted for in our models; we may yet have biases in our estimated dates. In addition, there are puzzling findings that need additional genetic data to resolve.
A trans-Pacific migration from southeast Asia does not in fact comport with the genetic evidence
One major unsolved mystery that whole genomes revealed is the shared ancestry between some South American and Australasian populations (from Australia, Melanesia and southeast Asia). Geneticists refer to this ancestry as from the ‘Ypikuéra population’ or ‘Population Y’. Population Y is seen scattered inconsistently in genomes throughout the Amazonian and Pacific coastal regions; it has been found in South American genomes as early as 10,000 years ago.
What could account for this pattern? It’s one of the biggest mysteries currently in the field. A trans-Pacific migration from southeast Asia that seems to offer an easy explanation does not in fact comport with the genetic evidence. Such a migration would leave a very different pattern of genomic footprints; Population Y ancestry is too old, too scarce, and too inconsistent to be explained by this model. Instead, we can trace the Population Y ancestry, tentatively, to East Asia; a 40,000-year-old individual from Tianyuan Cave in China carries its genetic signature. Most likely he represents a population no longer present in the region, which gave rise to both the ancestors of Australasians and Population Y.
So how did Population Y get into the Americas? Given current data, two scenarios suggest themselves. First, Population Y may have been present in the isolated group that gave rise to the different branches of the First Peoples during the LGM. It’s easy to imagine a scenario of a geographically dispersed metapopulation, consisting of multiple groups living in different refugia across Beringia, containing some families with this ancestry that simply didn’t get widely shared due to limited contact. If these different groups entered the Americas separately after 17,000 years ago, Population Y ancestry may have been limited to certain descendant populations.
Another possibility ties together evidence of Population Y and potential archaeological evidence at sites like White Sands. Could there have been multiple migrations into the Americas, with one pre-LGM migration consisting of Population Y individuals (and perhaps Unsampled Population A or other groups we have yet to identify), and one post-LGM migration? This would reconcile the archaeological evidence of early traces of humans in the Americas (if they are indeed legitimate, which has yet to be demonstrated to everyone’s satisfaction) and the genetic data.
Archaeologists sceptical of White Sands’ early dates are dubious of the second scenario, and it is admittedly speculative. We need a great deal more data, both genomic and archaeological, to test it. But, as a field, we are actively engaged in collecting that data, even as I type these words.
We may never know exactly how the White Sands people fit into the biological history of the Americas. But to members of the Pueblo of Acoma, near the White Sands site, the identity of these individuals is not at all mysterious. ‘Thousands and thousands of years ago, our ancestors walked this place,’ said Kim Charlie, a member of the Pueblo of Acoma, in an interview with Lizzie Wade for Science magazine in 2021. Their language even has words for the extinct megafauna seen at the site. In an example of how the discipline of archaeology is evolving for the better, the Pueblo of Acoma is actively involved in the recovery and study of these ancient footprints.
The addition of more ancient genomes from Indigenous populations across time and space will provide fascinating details about the lives, choices and movements of the earliest peoples of the Americas. We already see this occurring, with recently sequenced genomes from Brazil giving insights into migrations through South America. I am excited to see what the future reveals about the past. But I will venture to predict that, whatever my field does reveal, it will ultimately only affirm what Indigenous peoples already know to be true: they have been here since time immemorial.





